Final Thoughts from Newsletter III
A More General Critique
EPONA was also fortunate to receive another perceptive letter from Paul Preuss, the second in a row to be published so, even though he wasn't at CONTACT X, we can call him a Coti Mundi veteran.
"With due appreciation for the fascinating, creative thinking that has gone into Coti Mundi's plants and animals so far (by the way, I do like the name Epona better - Earth parochial perhaps, but not an awkward pun), I admit to some discouragement that what seems like a wonderful opportunity was let slip.
"Here we have a planet covered with water, in which the timeframe for evolution on land is intermittent and short, and yet the assumption seems to be that the interesting evolution will nevertheless happen on land and that our eventual sophont will be a landbased critter. At least that's where most of the work has gone so far. Why?"
Good questions Paul, but hopefully this issue puts some of your concerns to rest.
"Reading the precis, I confess I got more enjoyment out of Act I, where the concentration was on the ecosystem of the lagoons of the Sunken Continent, than on the landforms of Act 2, interesting and remarkable as they are. What is it about land, as opposed to protected or even open sea, that favours the evolution of an intelligent species? A propos flora, you correctly note that seaweed doesn't evolve to redwoods in 5 million years; as for intelligence, the only case on record indicates that it took more than 60 million years for humans to evolve once mammals got their chance. And what really gave us a distinctive edge was not so much our brain as our hand, which presumably required our primate ancestors to have spent lots of time in trees.
"Poncedonus is the presumed ancestor of whatever sophonts will evolve on the land surface of the Sunken Continent, a creature equipped with manipulative organs which its descendants seem to have lost rather than elaborated. (Certainly none mentioned so far have spent time in tress, which have not been invented yet.)
"Now there are bright and highly "manipulative" (telling etymology!) species in the seas of Earth, namely cephalopods, which on a different evolutionary track might well have outshone us primates in several measures of intelligence. (Many species of these, ammonites, were wiped out in the KT boundary event that coincidentally allowed mammals to come to the fore.) When it comes to intelligent life in the seas, we're not stuck with the sea otter/dolphin (i.e. readapted land creature) paradigm.
"This broadside is meant to propose that, when the time comes to think of a second sophont, instead of looking in the raw volcanic Highlands or on Fire Island, we look to the seas. Moreover, we ought to look there for an ancestor of Poncedonus (agreed, an awful name) to provide the common link.
"Meanwhile, evolution is opportunistic, and here are these three-limbed critters springing about on the dry land of the Sunken Continent. From the history of life on Earth, we see that several body plans may exist simultaneously (four limbed, six-limbed, eight-limbed, no-limbed etc.), but will occupy separate sets of niches (insects dominate absolutely, but vertebrates, most with four limbs, dominate where large size is a plus). Other planets, other niches. On Epona, three limbs might serve over a very wide range."
The Poncedonus clade is actually five limbed, although its Ceretridon descendants are effectively three-limbed because limbs 1&2 have become mouthparts.
"But we might profit by not thinking of the three-limbed critters of Epona as three-limbed; instead, let's think of them as tetrahedrons. Imagine carbon in a diamond configuration, a net of tetrahedrons, every atom equidistant from each other, a centre with four corners, to wit:"
Paul's sketch does in fact show five limbs - but the first two are small and close to the head.
"Thus, since we have to make our skeletons from calcium carbonate (limestone! - but it's a low gravity planet), the most important skeletal unit is the "pelvis", which occupies the creature's center. From this centre, jointed bony processes extend more or less equally up the "neck", down the "legs" and down the "tail". The terminal nodes might form new tetrahedrons, as in the diamond network. At the ends of the limbs (and tail), these nodes might sprout three toes and fingers; at the end of the neck, mouthparts and/or sense organs.
"The brain of the basic critter may be housed near the "pelvis" - assuming there is an ancestor, more basic than the Poncedonus, which operated not in a straight ahead, two dimensional way but in all three dimensions. (The analogy is to the octopus. We humans probably owe the location of our brains at the top end of our spinal cords to our noses, which we inherited from those blind, mud-sucking worms that were our ancestors. Since it was the nose that distinguished food from garbage, it functioned best on the leading end.) If the skull doesn't house the brain, it need only be a scaffold for muscles and a conduit for nerves - a very Swiss cheese-like affair. The pelvis, on the other hand, could be very interesting.
"As to the asshole problem, there is none in the basic structure: dumping takes place in the centre. In Ceretridon, sideways venting may have evolved as an ad hoc solution, just as migrating eyes evolved in flatfish. (The unrelated Springcroc probably doesn't have an anus; like a sea urchin, it ejects waste from the same opening that serves as its mouth.)
"Small tetrahedrons might take to the trees, the rocks (presumably the mountains here are especially rugged because of low gravity - and, especially if limestone, subject to fantastic sculpting by wind and water), and the air. Membranes between processes - say the neck and forelimbs, with the tail as stabilizer - would give us wings. It is from one of these agile little ones, rather from some awkward, mastodon-like three-legged plains grazer, that we might expect a sophont to descend."
Miscellany:
"Muscles extending, rather than contracting: add big cantilevers opposite all joints. A cute idea, but any payoff? Any evolutionary advantage over contracting muscles? None apparent to me. I don't think the creatures we've seen so far would have evolved at all, if muscle tissue was extensively extensile. We'd have a whole different set of beasts."
Paul Birch has dealt with this issue well and has sidestepped most of the problems. However, we will have to get our artists to redraw some of our creatures slightly to remove those Earth-like joints. This should please Jack Cohen at least who hates vertebrate type jointing, thinking it to be too parochial to Earth. Paul wrote:
"Homeostasis: I favour warmblooded creatures with "fur", not I hope for mammalian chauvinistic reasons but because fur is so versatile. Protects against the cold ¾ polar bears, seals, walruses etc. (but not Cetaceans, I grant you) - but adapts to climate change throughout shedding, changing colour etc., (manes, tails, spots, stripes, etc.). But that's a vote, not a reasoned argument.
“Plants, trees: the variety on Earth is so enormous I would only caution against too conservative an approach. By all means let's take the bubble weed as far as it will go, but even in a relatively short 10 million years there will be thousands of other inputs. Virtually any flora we want, we can have, consistent with red wavelength photosynthesis and low gravity."
Might We Have Aerial Plants?
Don Scott sent in this email:
"Could not plants which have developed the ability to float on water (like our algae with bladders) not extend that capability to floating in the atmosphere? Would the atmosphere on "our" world permit that? How would the intervening steps work? Where would the nutrients come from? Would they sink at night, like the Martian balloon, to gain nutrients from the sea, then rise as the sun warmed them? Then the day-rise of plants would protect them from the herbivores (or plant predating life forms).
"Would the animals then not evolve to pursue the new atmospheric plants? Perhaps the Springcroc or a relative might develop the ability to spring on a few plants. Or the skimmers might evolve their webbed feet, into some type of wing.
"The more I think about it, the more I like the concept of a "ballooner" plant. This is a very plausible evolution from the wandering (pelagic) plants which have already been described. This also allows an organism to develop which can take advantage of both ice and fire eras - in the sea during ice, on the land during fire. This plant - if it does descend from the "bubble" plant - does have some interesting evolutionary possibilities. Since the bubble plant is an evolutionary opportunist, capable of "explosive growth" in the right niche, and if we extend its
range by a floating-through-the-atmosphere capability, it has both qualities for high evolution: it can endure through many outside changes; it can explosively grow and develop when conditions are good; since evolution will occur more rapidly when growth is explosive, we can project some interesting evolutionary changes for this plant. Its dangling roots could become (on land) stronger supports; or even flexible, allowing the plant to move down close to the surface or high above it.
"Other creatures -- faunal -- could adapt to pursue the balloon plant on land. This might lead to longer necks, the ability to spring up to "capture" the plant, etc. But I still also think that such a plant could become at least a primitive sophont. If we factor in carnivorous qualities, for example, so that it hunts as well as gets hunted. Or if we really rethink the development of intelligence along new lines."
Keep going Don, it's a fascinating concept. I've often wondered why aerial plants have never evolved on Earth. Is it because it never happened, or because there is some insurmountable barrier to it being achievable? Biology has the demonstrated ability to produce hydrogen or methane gases which could provide the buoyancy and floating plants could get above the forest canopy to intercept the sunlight first. The mineral nutrients issue though is a problem, they would probably need to sink to the ground (probably when the gas cools at night) to obtain phosphorous, etc. Nitrogen needn't be a problem though, as we might conjecture the presence of nitrogen fixing symbiotes that can obtain N straight from the atmosphere.
Moore's Non-Carbon Photosynthesis
We would also like to thank David Moore for his late submission of a massive 19 page manuscript which considers a possible biochemical adaptations of Eponan plants to low levels of CO2. About half of the text is a detailed description of the nitrogen cycle which can be gleaned from any good undergraduate level biochemistry textbook and some of David's arguments are based on the erroneous assumption that all multicellular Eponan life becomes extinct. This only applied to the land surfaces. For these reasons, Greg and I have decided not to mail this manuscript complete with Newsletter #3 but to reflect on its contents at greater length and comment on them in a future issue.
However, David has come up with one fascinating idea which may be worth pursuing in the interim. Since plants have had to adapt to lower CO2 levels, he proposed an evolutionary innovation whereby photosynthesis might use non-carbon compounds for energy storage. David proposes nitrogen compounds and hence nitrogen cycling plants. The pages of equations and diagrams he uses to support the feasibility of this idea as a biochemical cycle, can be summarized in two simpler equations:
1) Photosynthesis: 3 H2O + N2 > 2 NH3 + 1.5 O2.
Delta G = +679.71 kJ/mole N2.
2) Respiration: 2 NH3 + 1.5 O2 > 3 H2O + N2.
Delta G = 679.71 kJ/mole N2.
Of course, many intermediate compounds are used such as nitrite and nitrate, etc., and carbon has to be fixed non-photosynthetically. David postulates the use of biomolecules low in carbon content to reduce reliance on this limiting substance.
There may be some profit in looking at this issue further although David's proposal that nitrogen oxides will build up in the atmosphere of Epona during Epochal Winters seems infeasible for a number of reasons that will have to be discussed properly in a future issue.
Revising the Family Tree
There is sufficient fresh material to permit the drawing of new evolutionary trees for Eponan creatures and this I have attempted in the next two figures.
Birch's suggestion that Pagoda trees and Pentapods share a related musculature is fascinating as it indicates that some Eponan plants and animals may not be nearly as distantly related as on Earth, where the Plant and Animal Kingdoms diverged at the unicellular stage. The fact that myoskeletal animals differ so fundamentally (on almost every anatomical and physiological count) from other animals, makes it clear that myoskeletal plants are closer relatives! Thus we cannot have a simple Plant/Animal Kingdom dichotomy as on Earth. This may not be as daft as it sounds because, although Epona once had a stable climate like Earth, the Epochal Summers and Winters into which it has now fallen have caused a succession of many severe mass extinctions. Almost any vestiges of the old order have therefore long been swept away.
I propose therefore a three Kingdom Phylogenetic system (not including bacteria).
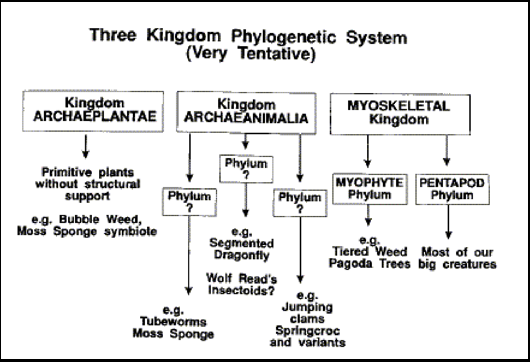
The ARCHAEPLANTAE were the plants that used to rule the world, but many of its former advanced forms are long extinct. For instance, if ancient plants one had the ability to make "wood", this has been lost. Archaeplantae now are "algae" types, such as aquatic "phytoplankton", mats of photosynthetic slime and symbiotic forms such as that which grows with the Moss Sponge. Some larger plants are still extant, but have only primitive structural support and on land therefore tend to grow as a moss-like coating on the ground rather than as a raised canopy. Once of the most successful and hardy of the Archaeplantae is the Bubble Weed.
The ARCHAEANIMALIA are animals with mineralized endo or exoskeletons (mostly carbonate) and tensile muscles. Again, they are much reduced from their former diversity ~1.7 billion years ago. Many groups are primitive and sessile, such as the reef building Tubeworms, the Moss Sponge and the host of bottom dwelling detritus feeders on the sea floor. One Phylum of creatures are moderately similar in form and function to terrestrial arthropods and comprise creatures such as the Segmented Dragonfly and possibly Wolf Read's insectoids. A bivalved, basket-like, endoskeleton is characteristic of the phylum that contains the Springcrocs. Members of this clade range from tiny filter feeding "sea shells" to the most massive animals on the planet.
The MYOSKELETAL Kingdom is a relatively new innovation in Epona, but is now a major taxonomic group and the one containing the most sophisticated forms. Its origin probably dates to not more than 2 billion years ago and may be as close as 1 billion. The progenitor was probably an Archaeplant that evolved primitive osmotic muscle that later become animal myoskeleton. MYOPHYTES (such as Tiered Seaweed and Pagoda Trees) and PENTAPODS are thus two Phyla in the same Kingdom, but with a very remote common ancestor. Myophytes now predominate as upstanding plants because of their ability to support themselves. Pentapods went on to evolve boost metabolism, giving them an advantage in many mobile, energy demanding, ecological niches. It is possible to imagine other myoskeletal phyla with differing body plans, but as yet we have no candidates.
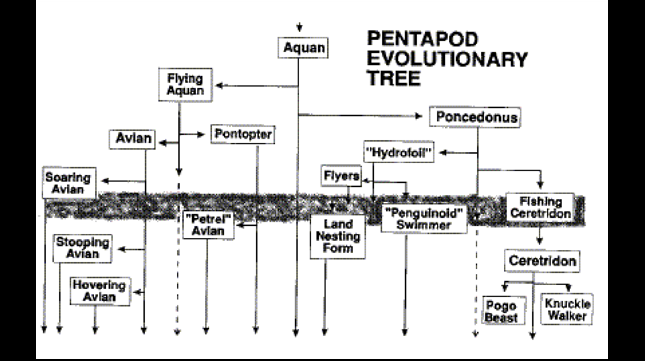
Of the Pentapods, Paul Birch has provided two new major classes -- the Aquans and the Avians -- and an intermediate link, the Flying Aquan. Del Cotter has evolved Pontopter from the Flying Aquan. We are therefore in a position to substantially augment Geoff Landis' Poncedonus evolutionary tree (Figure 4 of Newsletter #2), and we show my concept of a Pentapod tree that begins with the Aquan.
It's still tentative and will certainly be subject to revision as the creative process further unfolds. The horizontal grey band is the approximate time at which the Ice Epoch ends and dotted arrows represent my speculation of the two forms most likely to have become extinct. (Please note that there is one error in that the land herbivores, descended from the Poncedonus via the Snow Skimmer, have been inadvertently omitted.) It's evident therefore from this Figure that we now have considerable scope to advance towards an even more convincing fauna and ultimately, a sophont.
Just before going to press with this issue, Del Cotter submitted a taxonomy of Epona that supplements the work done by Martyn in classification. It is reproduced here complete with Del's comments following:
| Building a Taxonomy for Epona based on Earth Taxonomy examples | ||||
|---|---|---|---|---|
| Category | Suffix | Taxa | ||
| Example for a terrestrial plantPlants | ||||
| Kingdom | Plantae | |||
| Division | -phyta | Trachaeophyta | ||
| Class | -opsida | Magnoliopsida | ||
| Order | -ales | Asterales | ||
| Family | -acae | Asteracae | ||
| Genus | Bellis | |||
| Species | B. perennis | |||
| Examples for terrestrial animals | ||||
| Name | Human | Lion | Housefly | |
| Kingdom | Animalia | Animalia | Animalia | |
| Phylum | -a | Chordata | Chordata | Arthropoda |
| Class | -a, -ia | Mammalia | Mammalia | Insecta |
| Order | -a, iformes | Primata | Carnivora | Diptera |
| Family | -idae | Hominidae | Felidae | Muscidae |
| Genus | Homo | Felis | Musca | |
| Species | H. sapiens | F. leo | M. domestica | |
| Epona Taxonomy | ||||
| Name | Pagoda Tree | Mariner | Stooping Avian | |
| Kingdom | Myoskeleta | Myoskeleta | Myoskeleta | |
| Phylum | Myophyta | Pentapoda | Pentapoda | |
| Class | Tieropsida | Aquania | Aquania | |
| Order | Pagodales | Pontoptera | Deltiformes | |
| Family | Pagodacae | Marineridae | Raptoridae | |
type specimen: Pagoda birchii
and location: (pg. 1, Birch manuscript)
Del notes:
"I haven't suggested taxa right down to species, since I like to imagine that every sketch represents a different species, and every artist brings to life a new genus. Thus, although P. birchii will never be sketched by anyone else, other artists will sketch other members of the family or order of which it is the type. You can also see that I disagree with your characterization of Birch's avians as a genus; I see them as an order. We do not have a sufficiently detailed "telescope" with which to split taxonomic hairs so fine as to talk about genera. Consider the difficulty with which an alien would have distinguishing different members of the family Hominidae!"