Inside the Pentapods
Musculature & Respiratory Systems
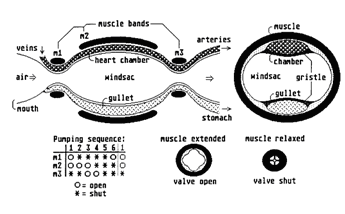
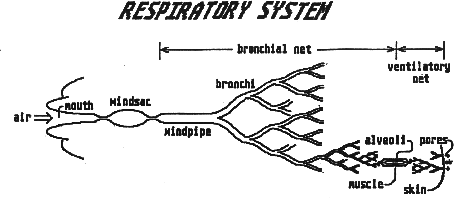
Another useful contribution Paul has made is to provide details of the internal anatomy and physiology of his Avians. Since these creatures share a common ancestor with Poncedonus, then many of these features will be basically the same for all members of what we might call the "Pentapod Phylum". The muscular combination windsac/heart/gullet will therefore be similar for both Avian and Ceretridon; as will be the ingenious way air is passed directly through the body and is expired through pores in the skin.
Reading page 3 of Paul's document, a number of implications instantly spring to mind: For a start, the use of skin pores to exhale both respired air and water vapor coolant, immediately places constraints on skin coverings. Thick fur, impervious scales, or viscid mucous do not seem advisable. Creatures instead may adopt other methods of protection from cold, or maybe their ferocious metabolic rate makes hypothermia less of a problem.
The need to rhythmically contract the windsac, has particularly interesting connotations for the production of sound. Pentapod "speech" will have a continuous background beat to it, like that of a disco record. Feel free, any of you, to think up other logical implications of this internal design . . .
At CONTACT X, nothing was done about any of our creatures' skeletal support, (except for the Springcroc which is not a member of the Pentapod Phylum and may not even be in the same taxonomic Kingdom -- see later). Geoff Landis' suggestion of exploring the implications of extensile muscles was mentioned literally in the last 30 seconds of the conference. Since the whole point of the Coti Mundi Project is to go beyond cardboard cutout aliens that are little more than works of art to be looked at, then such considerations are essential.
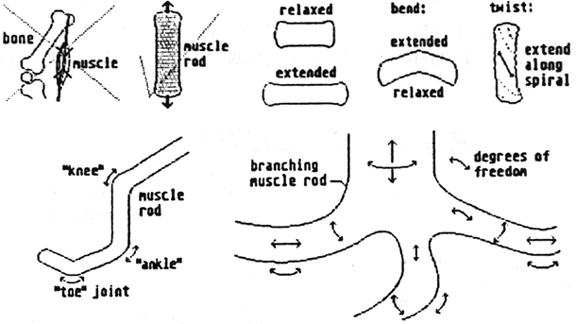
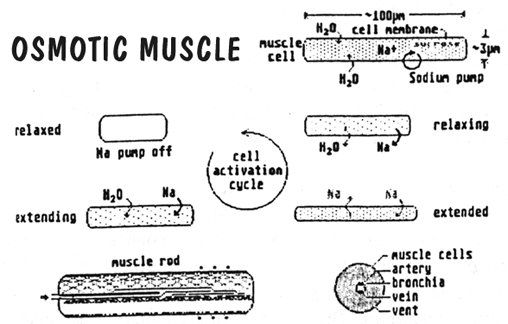
Again Paul has provided us with both an extremely elegant solution and a wonderful challenge. He has provided a good rationale for extensile muscle rods that actually make a mineralized skeleton redundant. In fact the function of bones and muscles are combined (as a "myoskeleton") in a way such that some aspects of mobility are better than a terrestrial vertebrate's jointed skeleton and some not so. This forces us to think differently, pushing us down a different and more creative logical path. Plausible inventions like this are precisely what a good alien design undertaking needs.
Birch's text adequately explains the operation of his osmotic myoskeleton, along with its ability to bend and twist. Obvious implications of this are that Pentapods will have no true joints in the sense that we understand them on Earth. Thick muscle rods however will probably not be very flexible and will certainly not bend through such a sharp angle. However, flexure will not be restricted to one junction and therefore a limb will tend to bow in a curve, rather than folding. It is possible that certain limbs may not need to be equally flexible everywhere and efficient locomotion might require bending to occur in one preferred location. In which case, this section of the muscle rod could become more vascular and specialized for flexure, forming a "pseudojoint". Thinner, less stiff muscle rods will be more bendy and therefore Pentapod manipulanda will be more like short tentacles than our bony fingers. Static myoskeleton, without extensile ability, and which is denser and more cartilaginous, might act as a superstructure or protective casing for vulnerable internal organs.
Interesting, eh? This is certainly something that can be taken further. For instance, I'm slowly putting together a generic design for Pentapod (preCeretridonal) mouthparts. How about "chitinous teeth" with a deep, screw-like thread attached to extensile myoskeletal rods? As the teeth are thrust into the victim, they also twist and lock . . . watch this space . . .
One criticism of the myoskeleton might be that our artists will have to redraw all the pictures that they have done so far. Not so. Look back at Marc Moellinger's rendering of Ceretridon in Act II -- the joints are not clearly defined here and could just as well be "pseudo joints" -- and this is the best picture we've got. Anyway, I'm not proposing all our creatures should be myoskeletal, only those of the Pentapod Phylum and relatives (see later). Springcroc and a number of other beasts from the lagoon stage use mineralized skeletons and probably more Earth-like, tensile muscles.
The Ability to “Kick Ass”
Last issue, I posed the possibility -- half in jest -- of endowing some of our organisms with a beefed up metabolism that, ". . . turns the creature into something that, just for short while, could turn Arnie Schwartzenegger into shish kebab!"

Paul also decided to tackle this issue, not by using nitrous oxide, as I suggested, but hydrogen peroxide oxidizer and ethanol "fuel". The final page of his submission therefore presents a scheme called "muscle boost" which permits myosketetal animals to operate with, "...the roughly tenfold gain in muscle power it affords." This is particularly effective for flying creatures, giving them the ability to perform long, high speed journeys.
This muscle boost idea is bound to be the most controversial of those Paul has had to offer, especially as it is the least clearly explained. A lot of what I have to say, therefore, has come out of further exploration of the concept over the telephone. His equations check out, but, in themselves, do not give anything like the augmented power output above ordinary glucose -oxygen respiration that is claimed. This is because Birch is also exploiting the fact that he is using a liquid oxidizer, H2O2, which can be supplied at a much greater rate than haemoglobin in terrestrial blood can provide oxygen. "Boosting" cannot be switched on instantly and requires the secretion of a hormonal trigger, either consciously, or in response to the sort of "fight flight and fright" reflex that triggers mammalian adrenal glands. Internal sacs that store ethanol and H2O2 (separately) then release these substances into the blood stream. Because H2O2 is so damaging to cytoplasm (especially in high concentrations, it is only taken up into muscle cells at the rate it is being consumed, so the storage sacs, after their initial contraction, keep the oxidizer concentration topped up at ~1%. Blood vessels, therefore, are lined with material that is resistant to peroxide, keeping it confined (except in very low concentrations) to the storage and circulatory systems only.
Unlike the Niven/Pournelle/Barnes version of a super metabolism in Legacy of Heorot, Birch's boost is not for augmenting modes of behavior where brief surges of power are needed. The efficient water vapor/air cooling system allows metabolic augmentation to be used for extensive periods of up to several hours (as in the case of a full-charged animal that has prepared itself for migration). It endows not so much super-strength (since this is dependent on muscle mass) as a phenomenal endurance. This permits avians to attain high speeds (several hundred m.p.h) and therefore to travel long distances without tiring in a comparatively short period of time. Landbased creatures do not gain so much in speed (due to difficulties with traction), but can engage in indomitable struggle for as long as the stored metabolites last. A migrating herd of herbivores might negotiate the steepest mountain pass in an afternoon, whilst an army of sophonts might fight with incredible stamina, every blow a haymaker, and the last strike being as fresh as the first! Of course, when boost finally runs out, fatigue sets in as fast as in humans and the first side in a long battle to reach exhaustion will lose. Of course, once technology is invented, devices to inject extra peroxide and ethanol could arbitrarily increase the time spent boosting -- we could perhaps have a clan, or social class of short-lived, hard fighting, Viking-type super-beserkers! A brief lifespan would not just result from the high chance of being a casualty in battle, but also from the more insidious effects of excessive internal oxidation, resulting in accelerated aging and an even greater death wish.

This metabolic option does have its drawbacks however. An obvious one is that more food must be consumed as it is being stored in a concentrated form that can be used and depleted rapidly. If we propose that animals only start synthesizing new stores of ethanol and peroxide when they are resting, then an increased requirement for some sort of "sleep" may be indicated. Tired creatures on a migration, or a forced march after a battle, would therefore be particularly vulnerable to attack by rested enemies. Recovery from serious exhaustion, involving a substantial loss of body weight, could take many days of heavy feeding. More modest use of boost metabolites, however, should prevent storage sacs from emptying entirely, so that a reserve is kept available to deal with random events.
Another difficulty is that boost cannot be "switched off" nearly as fast as it can be "turned on". This is because the H2O2 concentration is maintained at ~1% in the blood; once the hormonal signal arrives at the storage sacs to stop any further release, there is still a large, unwanted excess sloshing about in the vascular system. This will have to be metabolized and so hyperactivity will continue -- the creature may take up to an hour to "wind down."
It is clear, therefore, that endowing some of our creatures with such a metabolic trick, increases our opportunity to be creative in some exciting and unexpected directions. The potential implications of boosting will be great and will require a good deal of thinking through. But then again, that's what we should be all about.
A NOTE ON THE SPRINGCROC
Pentapods are not entirely dominant as the large animals in land habitats as the ability to boost does not confer any advantages to reaction time. Eponan predators specialized for ambush are therefore non-myoskeletal creatures that rely on surprise and an ability to incapacitate a victim in less than two or three seconds. Springcrocs are of this type which, as was noted in Newsletter #2, lie in wait for victims, in pools or camouflaged as rocks, and engulf prey whole in an extensible stomach. Small and mid sized Pentapods are the usual food and are killed by being stung and injected with an enzyme such as catalase. As the victim boosts to try and escape, H2O2 in the vascular system violently dissociates into oxygen causing massive shock, almost instant incapacity and death shortly afterwards. Since Springcrocs are not designed for long distance locomotion and do not possess boost, they cannot compete with Pentapods in other niches. However, they do very well in their own, due to sheer size, bulk muscle power and calcite/aragonite armor plating just under the skin. Once their upper and lower body shells close, engulfed victims are doomed and the Springcroc itself is almost as invulnerable as an Ankylosaurus.

We could do with some development of this clade too. I don't feel they are slated for sentience, but one can imagine a number of nasty variants on the basic form; giants perhaps, or ones where the poisonous spines fringing the shell edges have evolved to cover the upper half of the body. Artists are also invited to illustrate this most evil looking of Epona's creatures.
Ready to find out more?
The curtain falls